 Our research seeks to understand the fundamental, yet long mysterious, cellular mechanisms that drive chromosome dynamics during the differentiation of sex cells (gametes).
Our research seeks to understand the fundamental, yet long mysterious, cellular mechanisms that drive chromosome dynamics during the differentiation of sex cells (gametes).
A critical feature of gamete differentiation is chromosome reduction; each gamete must contain exactly half the chromosome complement of its progenitor (parent) cell. If chromosomes fail to segregate properly during gamete formation, gametes and the offspring they generate are aneuploid (contain an improper number of chromosomes).
How are chromosome complements properly divided? At the onset of the specialized cell division cycle called meiosis, each chromosome somehow identifies and specifically associates with its homologous partner. This nuclear reorganization process culminates in paired homologous chromosomes that are joined along their lengths by a proteinaceous structure, the synaptonemal complex (SC), and each capable of undergoing crossover recombination. Each of these steps, chromosome pairing, SC assembly (synapsis) and crossover recombination, are conserved features of meiosis that ensure accurate chromosome reduction: Together, these steps allow homologous chromosomes to orient with respect to one another and thereby segregate toward opposite spindle poles at the first meiotic division. Despite over a century of observing meiotic chromosome pairing and synapsis in diverse organisms, the molecular mechanisms that promote and coordinate these fundamental meiotic chromosomal events are still unknown. How do homologous chromosomes identify one another? How is this initial recognition between chromosomes reinforced? How is homolog recognition coordinated with SC assembly, such that synapsis occurs specifically between paired chromosomes?
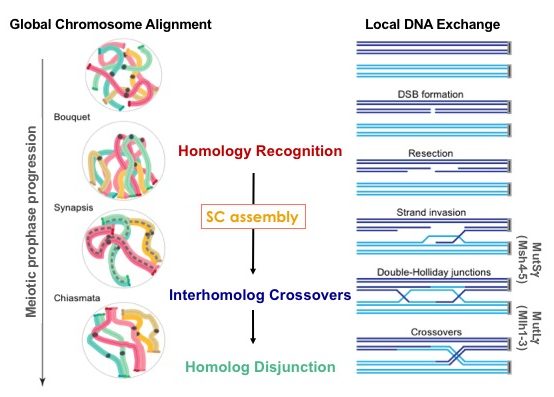 Our research is currently aimed at obtaining a molecular view of SC architecture and of the functional relationship between the structural building blocks of SC (particularly the SC transverse filament protein Zip1) and meiotic crossover recombination in budding yeast. Zip1 is predicted to assemble dimeric rod-shaped units that we know assemble with their C termini oriented toward homolog axes and their N termini oriented toward the midline of the SC. Our lab discovered that the Ecm11 and Gmc2 proteins assemble the central element of the budding yeast SC. This work also brought us to the discovery that, although crossover recombination events take place in the context of the SC, mature SC structure is dispensable for crossover recombination in budding yeast: ecm11 and gmc2 null mutants fail to assemble mature SC but exhibit robust (even excess) crossover recombination.
Our research is currently aimed at obtaining a molecular view of SC architecture and of the functional relationship between the structural building blocks of SC (particularly the SC transverse filament protein Zip1) and meiotic crossover recombination in budding yeast. Zip1 is predicted to assemble dimeric rod-shaped units that we know assemble with their C termini oriented toward homolog axes and their N termini oriented toward the midline of the SC. Our lab discovered that the Ecm11 and Gmc2 proteins assemble the central element of the budding yeast SC. This work also brought us to the discovery that, although crossover recombination events take place in the context of the SC, mature SC structure is dispensable for crossover recombination in budding yeast: ecm11 and gmc2 null mutants fail to assemble mature SC but exhibit robust (even excess) crossover recombination.

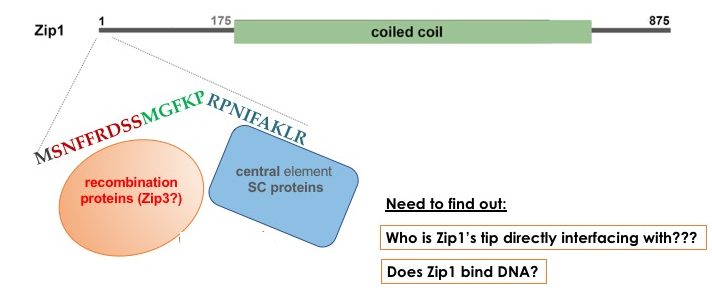
 A focus that underlies many of our recent experiments has been to identify molecular partners and features of the SC transverse filament protein Zip1 that underlie its crossover-promoting and SC assembly activities. Our work in this area has demonstrated unequivocally that Zip1’s SC assembly and crossover recombination functions are independent. Furthermore, our data suggests the Zip1 coordinates its crossover-promoting and SC assembly activities through adjacent small regions within its N terminal tip.
A focus that underlies many of our recent experiments has been to identify molecular partners and features of the SC transverse filament protein Zip1 that underlie its crossover-promoting and SC assembly activities. Our work in this area has demonstrated unequivocally that Zip1’s SC assembly and crossover recombination functions are independent. Furthermore, our data suggests the Zip1 coordinates its crossover-promoting and SC assembly activities through adjacent small regions within its N terminal tip.
We have furthermore established fruitful collaborations with Dr. Owen Davies at Newcastle University, and with Dr. Ishita Mukerji in our own department, to begin analyzing the structural properties of SC proteins, including Zip1 and the Ecm11-Gmc2 heterocomplex.
Our careful phenotypic analysis of crossover recombination in a wide variety of meiotic mutants (primarily the hard work of a large number of undergraduate students over the past two to three years!) also revealed an unexpected link between structural proteins of the SC and mismatch repair, a process that is fundamental to most if not all interhomolog recombination events that occur during meiosis. In collaboration with Dr. Jennifer Fung (UCSF), we have generated genome-wide profiles of mismatch repair in the meiocytes of our mismatch repair-defective mutants. This dataset confirms our initial genetic evidence of mismatch repair errors in our mutants, and allows us to examine the profile of recombination events that suffer from defective mismatch repair.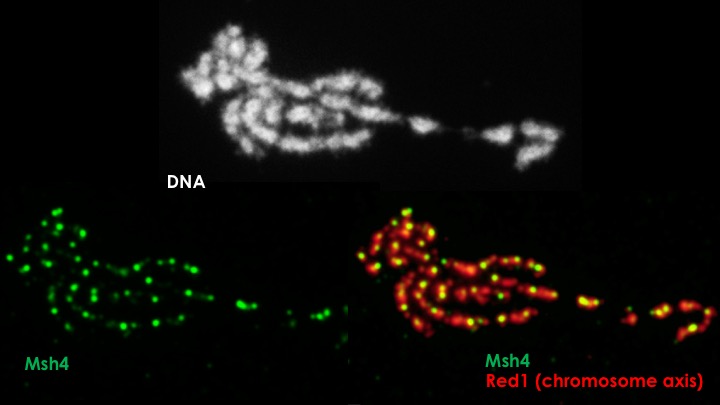
A fundamental goal of the lab’s current and future research is to understand how homologous chromosome pairing and SC assembly are coordinated with the meiotic recombination process in budding yeast.
Funding: National Institutes of Health
![]() Current lab members: Karen Voelkel-Meiman (research partner in crime), Chandni Ravindan (doctoral candidate), Sabrina Sharmin (doctoral candidate), Arpie Bakshian (rising senior), Gavin Shamis (rising junior), Charlotte George (rising senior).
Current lab members: Karen Voelkel-Meiman (research partner in crime), Chandni Ravindan (doctoral candidate), Sabrina Sharmin (doctoral candidate), Arpie Bakshian (rising senior), Gavin Shamis (rising junior), Charlotte George (rising senior).
